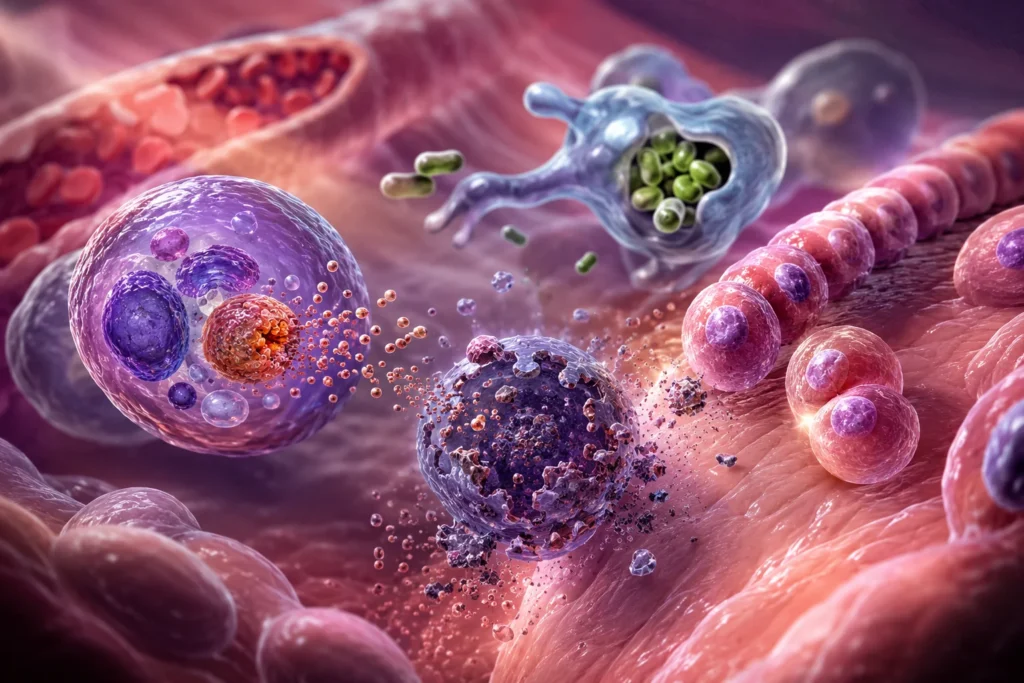
Los lisosomas constituyen uno de los sistemas fundamentales para el mantenimiento de la homeostasis tisular, la remodelación fisiológica de los órganos, la eliminación de estructuras celulares envejecidas y la destrucción de células irreversiblemente dañadas. Estas funciones dependen de su capacidad para contener un amplio repertorio de enzimas hidrolíticas capaces de degradar proteínas, lípidos, polisacáridos, ácidos nucleicos y otros componentes biológicos en un entorno altamente ácido. Gracias a estas propiedades, los lisosomas participan tanto en procesos fisiológicos normales de regresión tisular como en mecanismos patológicos relacionados con la muerte celular y la eliminación de microorganismos.
Los lisosomas son orgánulos rodeados por una membrana simple que contienen más de 60 enzimas hidrolíticas ácidas especializadas en la degradación intracelular de prácticamente todas las macromoléculas biológicas. Estas enzimas incluyen proteasas, nucleasas, glucosidasas, fosfolipasas, sulfatasas y lipasas. El funcionamiento óptimo de estas hidrolasas requiere un pH aproximado de 4.5 a 5.0, mantenido por bombas de protones dependientes de adenosín trifosfato localizadas en la membrana lisosomal. Esta acidificación permite una degradación eficiente de materiales intracelulares y extracelulares incorporados mediante diferentes mecanismos de transporte vesicular.
La capacidad degradativa de los lisosomas resulta esencial para la remodelación continua de los tejidos. En numerosos órganos, las necesidades funcionales cambian a lo largo de la vida o en respuesta a modificaciones fisiológicas, hormonales o ambientales. Cuando un tejido deja de requerir el volumen celular previamente alcanzado, se activan mecanismos de degradación intracelular que permiten reducir progresivamente su tamaño. Una parte importante de esta regresión depende de la actividad lisosomal y de los procesos de autofagia asociados.
La retracción tisular constituye un fenómeno fisiológico ampliamente distribuido en los organismos multicelulares. Durante este proceso, células previamente hipertrofiadas o altamente activas reducen su contenido citoplasmático mediante la degradación controlada de orgánulos y proteínas intracelulares. Los lisosomas actúan como centros de reciclaje donde estos componentes son descompuestos en moléculas simples reutilizables por la propia célula o por células vecinas.
Uno de los ejemplos más estudiados de retracción tisular ocurre en el útero después del embarazo. Durante la gestación, las células musculares lisas uterinas experimentan una marcada hipertrofia inducida por estímulos hormonales y mecánicos. Tras el parto, el volumen uterino disminuye de manera progresiva mediante un proceso denominado involución uterina. Durante esta regresión, una gran cantidad de proteínas contráctiles, mitocondrias y otros componentes citoplasmáticos son degradados por mecanismos autofágicos dependientes de lisosomas. La activación de la autofagia permite la eliminación ordenada de los elementos celulares excedentes y facilita el retorno del órgano a sus dimensiones previas al embarazo.
Procesos similares ocurren en el músculo esquelético durante períodos prolongados de inactividad. La inmovilización, la denervación, el reposo prolongado o la ausencia de carga mecánica inducen atrofia muscular. En estas circunstancias se incrementa significativamente la actividad de las vías autofágico-lisosomales. Los lisosomas degradan proteínas miofibrilares, orgánulos envejecidos y componentes citoplasmáticos que ya no son necesarios para mantener el volumen muscular previamente existente. La activación excesiva o prolongada de estas rutas puede contribuir a una pérdida importante de masa muscular.
Las glándulas mamarias proporcionan otro ejemplo clásico de regresión tisular dependiente de lisosomas. Durante la lactancia, las células epiteliales secretoras experimentan una intensa actividad biosintética para producir leche. Al finalizar la lactancia, la glándula mamaria inicia un proceso de involución caracterizado por apoptosis, autofagia y remodelación extracelular. Los lisosomas participan activamente en la degradación de organelos secretores, proteínas acumuladas y estructuras celulares que ya no son necesarias. Este fenómeno permite que la glándula recupere progresivamente una arquitectura semejante a la existente antes de la gestación.
La relación entre lisosomas y autofagia es fundamental para comprender estos procesos de regresión. La autofagia constituye un mecanismo altamente conservado mediante el cual las células encapsulan porciones de su propio citoplasma dentro de estructuras denominadas autofagosomas. Posteriormente, los autofagosomas se fusionan con lisosomas para formar autofagolisosomas, donde ocurre la degradación enzimática de los materiales capturados. Este proceso permite eliminar componentes dañados, reciclar nutrientes y adaptar el tamaño celular a nuevas condiciones fisiológicas.
Además de participar en la remodelación fisiológica, los lisosomas desempeñan un papel crucial en la respuesta frente al daño celular. Las células están constantemente expuestas a múltiples agentes lesivos, incluyendo temperaturas extremas, radiación, estrés oxidativo, traumatismos mecánicos, hipoxia, toxinas y productos químicos. Cuando estos factores producen alteraciones estructurales importantes, los lisosomas intervienen en la eliminación de los componentes afectados.
El daño celular puede comprometer la integridad de las membranas lisosomales. La permeabilización de estas membranas permite la liberación parcial o total de hidrolasas hacia el citoplasma. Dependiendo de la magnitud del daño, las consecuencias pueden variar desde una degradación localizada de componentes celulares hasta la destrucción completa de la célula.
Cuando la lesión es limitada, solamente una región específica de la célula resulta afectada. En estas circunstancias, las enzimas lisosomales degradan selectivamente las estructuras dañadas, permitiendo posteriormente su sustitución mediante procesos de reparación celular. Este mecanismo contribuye a preservar la viabilidad de la célula mientras elimina componentes potencialmente perjudiciales. La capacidad de realizar una degradación localizada constituye una estrategia eficiente para mantener la integridad funcional del tejido.
Sin embargo, cuando el daño supera la capacidad de reparación, la liberación masiva de enzimas lisosomales desencadena la digestión progresiva de componentes esenciales de la célula. Este fenómeno se conoce como autólisis. Durante la autólisis, las hidrolasas degradan proteínas estructurales, membranas, ácidos nucleicos y orgánulos intracelulares, conduciendo finalmente a la destrucción completa de la célula.
La autólisis representa un mecanismo importante para eliminar células que han sufrido daños irreversibles. Desde una perspectiva fisiológica, este proceso evita que células gravemente lesionadas permanezcan dentro de los tejidos y comprometan la función del órgano. Una vez eliminadas, las células perdidas suelen ser reemplazadas mediante proliferación mitótica de células vecinas o por activación de poblaciones de células madre residentes.
Actualmente se reconoce que la permeabilización de la membrana lisosomal constituye un punto crítico en la regulación de diversas formas de muerte celular. La liberación citoplasmática de catepsinas, especialmente catepsinas B, D y L, puede activar rutas apoptóticas, necróticas o autofágicas dependiendo del contexto biológico. Estas proteasas participan en complejas redes de señalización que conectan el daño lisosomal con la decisión celular de sobrevivir o morir.
Los lisosomas también poseen una función esencial en la defensa inmunológica frente a microorganismos invasores. Durante la fagocitosis, células especializadas como macrófagos y neutrófilos internalizan bacterias, hongos y otros patógenos dentro de vacuolas denominadas fagosomas. Posteriormente, los fagosomas se fusionan con lisosomas para formar fagolisosomas, estructuras altamente especializadas en la destrucción microbiana.
Dentro del fagolisosoma, los microorganismos quedan expuestos a un entorno extremadamente hostil. Uno de los principales mecanismos antimicrobianos es la acidificación del compartimento hasta valores cercanos a pH 5. Este ambiente ácido favorece la activación de las hidrolasas lisosomales y simultáneamente altera numerosos procesos metabólicos bacterianos esenciales para la supervivencia.
Entre las moléculas bactericidas presentes en los lisosomas destaca la lisozima. Esta enzima hidroliza enlaces específicos del peptidoglucano que constituye la pared celular de muchas bacterias. La degradación de esta estructura debilita la integridad mecánica bacteriana y favorece la lisis osmótica del microorganismo.
Otra molécula importante es la lactoferrina. Esta glicoproteína posee una elevada afinidad por el hierro, elemento indispensable para el crecimiento y metabolismo de numerosos microorganismos. Al secuestrar hierro libre, la lactoferrina genera un ambiente nutricionalmente desfavorable para las bacterias, limitando su proliferación y aumentando su susceptibilidad a otros mecanismos antimicrobianos.
Además de la lisozima y la lactoferrina, los lisosomas contienen múltiples péptidos antimicrobianos, proteasas, nucleasas y especies reactivas capaces de destruir microorganismos internalizados. La acción combinada de estos factores permite una eliminación eficiente de agentes infecciosos antes de que puedan causar daño significativo a la célula hospedadora.
Los avances recientes han demostrado que los lisosomas no son simples compartimentos digestivos, sino centros reguladores que coordinan metabolismo, crecimiento celular, respuesta al estrés, señalización inmunológica y supervivencia celular. Su participación simultánea en la remodelación tisular, la eliminación de células dañadas, la autofagia y la defensa antimicrobiana los convierte en elementos indispensables para el mantenimiento de la integridad estructural y funcional de los organismos multicelulares.


Fuente y lecturas recomendadas:
- Ballard, O. A., & Morrow, A. L. (2013). Human milk composition: Nutrients and bioactive factors. Pediatric Clinics of North America, 60(1), 49–74. https://doi.org/10.1016/j.pcl.2012.10.002
- Boya, P., González-Polo, R. A., Casares, N., Perfettini, J. L., Dessen, P., Larochette, N., Métivier, D., Meley, D., Souquère, S., Yoshimori, T., Pierron, G., Codogno, P., & Kroemer, G. (2005). Inhibition of macroautophagy triggers apoptosis. Molecular and Cellular Biology, 25(3), 1025–1040. https://doi.org/10.1128/MCB.25.3.1025-1040.2005
- Cuervo, A. M., & Dice, J. F. (2000). When lysosomes get old. Experimental Gerontology, 35(2), 119–131. https://doi.org/10.1016/S0531-5565(00)00088-9
- de Duve, C. (2005). The lysosome turns fifty. Nature Cell Biology, 7(9), 847–849. https://doi.org/10.1038/ncb0905-847
- Glick, D., Barth, S., & Macleod, K. F. (2010). Autophagy: Cellular and molecular mechanisms. The Journal of Pathology, 221(1), 3–12. https://doi.org/10.1002/path.2697
- Lawrence, T., & Natoli, G. (2011). Transcriptional regulation of macrophage polarization: Enabling diversity with identity. Nature Reviews Immunology, 11(11), 750–761. https://doi.org/10.1038/nri3088
- Mizushima, N., Levine, B., Cuervo, A. M., & Klionsky, D. J. (2008). Autophagy fights disease through cellular self-digestion. Nature, 451(7182), 1069–1075. https://doi.org/10.1038/nature06639
- Ohsumi, Y. (2014). Historical landmarks of autophagy research. Cell Research, 24(1), 9–23. https://doi.org/10.1038/cr.2013.169
- Settembre, C., Fraldi, A., Medina, D. L., & Ballabio, A. (2013). Signals from the lysosome: A control centre for cellular clearance and energy metabolism. Nature Reviews Molecular Cell Biology, 14(5), 283–296. https://doi.org/10.1038/nrm3565
- Turk, V., Stoka, V., Vasiljeva, O., Renko, M., Sun, T., Turk, B., & Turk, D. (2012). Cysteine cathepsins: From structure, function and regulation to new frontiers. Biochimica et Biophysica Acta, 1824(1), 68–88. https://doi.org/10.1016/j.bbapap.2011.10.002
- Wohlgemuth, S. E., Seo, A. Y., Marzetti, E., Lees, H. A., & Leeuwenburgh, C. (2010). Skeletal muscle autophagy and apoptosis during aging: Effects of calorie restriction and life-long exercise. Experimental Gerontology, 45(2), 138–148. https://doi.org/10.1016/j.exger.2009.11.002
- Yoshimori, T., & Amano, A. (2009). Group A Streptococcus: A loser in the battle with autophagy. Current Topics in Microbiology and Immunology, 335, 217–226. https://doi.org/10.1007/978-3-642-00302-8_11
Aprende administración paso a paso

ADMINISTRACION DESDE CERO