La pinocitosis es un mecanismo fundamental de transporte vesicular mediante el cual las células incorporan líquido extracelular y las sustancias disueltas en él a través de invaginaciones de la membrana plasmática que originan vesículas intracelulares. Este proceso constituye una forma especializada de endocitosis y se encuentra presente de manera continua en prácticamente todas las células eucariotas. Su importancia biológica radica en que permite la captación de nutrientes, proteínas, lípidos, hormonas, factores de crecimiento y otras macromoléculas que, debido a su tamaño o propiedades fisicoquímicas, no pueden atravesar directamente la bicapa lipídica mediante difusión simple o transporte mediado por canales.
La pinocitosis recibe frecuentemente la denominación de “bebida celular” porque implica la incorporación de pequeñas cantidades de líquido extracelular junto con las moléculas que contiene. Sin embargo, esta descripción simplificada no refleja la enorme complejidad molecular del proceso. En realidad, la pinocitosis representa un sistema altamente regulado que participa en la nutrición celular, la señalización intracelular, la regulación de receptores de membrana, la respuesta inmunitaria y el mantenimiento de la homeostasis tisular.
La actividad pinocítica ocurre de manera permanente en la mayoría de las células. No obstante, algunas presentan tasas particularmente elevadas debido a sus funciones fisiológicas especializadas. Los macrófagos constituyen uno de los ejemplos más destacados. Estas células inmunitarias poseen una intensa actividad endocítica que les permite explorar continuamente su entorno en busca de microorganismos, partículas extrañas, restos celulares y moléculas de señalización. Estudios cuantitativos han demostrado que los macrófagos pueden internalizar aproximadamente el 3 % de su superficie de membrana plasmática cada minuto mediante procesos endocíticos, lo que implica una constante renovación de la membrana celular y una intensa actividad vesicular.
Las vesículas originadas durante la pinocitosis suelen poseer diámetros comprendidos entre aproximadamente 100 y 200 nm. Debido a sus reducidas dimensiones, estas estructuras generalmente no pueden observarse mediante microscopía óptica convencional y requieren técnicas de microscopía electrónica para su visualización detallada. La introducción de la microscopía electrónica permitió identificar la existencia de depresiones especializadas en la superficie celular que participan activamente en la formación de estas vesículas.
La relevancia fisiológica de la pinocitosis se relaciona directamente con la necesidad celular de incorporar moléculas de gran tamaño. Las proteínas plasmáticas, por ejemplo, suelen presentar masas moleculares demasiado elevadas para atravesar espontáneamente la membrana plasmática. Como consecuencia, la pinocitosis constituye uno de los principales mecanismos que permiten su internalización. Este fenómeno resulta esencial para la captación de albúmina, transferrina, lipoproteínas de baja densidad, hormonas peptídicas, complejos inmunitarios y numerosos factores de crecimiento.
La velocidad de formación de vesículas pinocíticas aumenta considerablemente cuando determinadas macromoléculas se unen a receptores específicos localizados en la superficie celular. Este mecanismo recibe el nombre de endocitosis mediada por receptores y representa una forma altamente eficiente y selectiva de pinocitosis. Gracias a este sistema, las células pueden concentrar moléculas específicas del medio extracelular y transportarlas hacia el interior incluso cuando estas se encuentran presentes en concentraciones relativamente bajas.
La especificidad de este proceso depende de la existencia de receptores proteicos transmembrana especializados. Cada receptor reconoce ligandos concretos mediante interacciones moleculares altamente selectivas. Cuando una molécula extracelular se une a su receptor correspondiente, se desencadenan cambios conformacionales que activan la maquinaria molecular responsable de la formación de la vesícula. Esta selectividad permite a la célula discriminar entre diferentes moléculas presentes en su entorno y regular con precisión qué sustancias serán internalizadas.
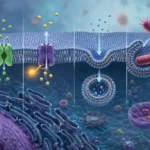
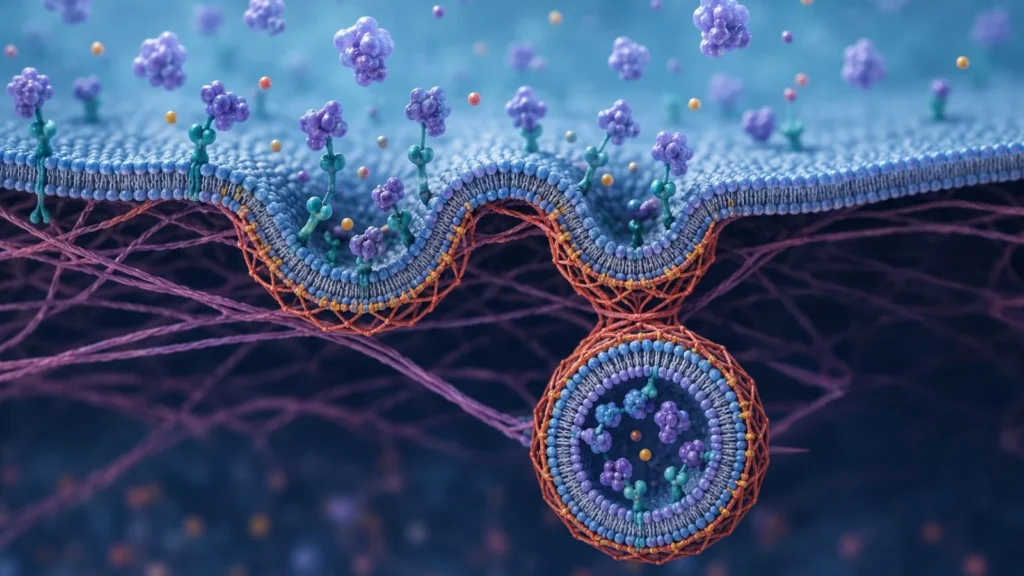
Los receptores implicados en la pinocitosis mediada por receptores no se distribuyen aleatoriamente sobre la membrana plasmática. Por el contrario, suelen concentrarse en regiones especializadas conocidas como hendiduras revestidas. Estas estructuras aparecen como pequeñas depresiones de la superficie celular y constituyen verdaderos centros organizadores de la endocitosis.
La característica más distintiva de las hendiduras revestidas es la presencia de una cubierta proteica formada principalmente por clatrina. La clatrina es una proteína estructural que se organiza en complejos tridimensionales denominados trisqueliones. Cada trisquelión está constituido por tres cadenas pesadas y tres cadenas ligeras que se ensamblan formando una estructura similar a una hélice de tres brazos. Cuando numerosos trisqueliones se unen entre sí, generan una red poliédrica que induce la curvatura progresiva de la membrana plasmática.
La formación de esta cubierta constituye uno de los eventos centrales de la pinocitosis. Diversas proteínas adaptadoras conectan los receptores transmembrana con la red de clatrina, permitiendo que las moléculas unidas a los receptores sean concentradas dentro de la región que dará origen a la futura vesícula. Este proceso incrementa notablemente la eficiencia de captación, ya que numerosas moléculas pueden ser internalizadas simultáneamente en una única vesícula.
Una vez que los ligandos extracelulares se han unido a sus receptores correspondientes, se producen modificaciones locales en las propiedades físicas y bioquímicas de la membrana. Estas modificaciones favorecen la invaginación progresiva de la hendidura revestida hacia el interior celular. A medida que la curvatura aumenta, la red de clatrina continúa ensamblándose y proporciona soporte mecánico para la deformación de la membrana.
Paralelamente, diversas proteínas accesorias participan en la regulación del proceso. Entre ellas destacan proteínas adaptadoras, proteínas que detectan la curvatura de la membrana y proteínas reguladoras del citoesqueleto. La coordinación entre estos componentes permite que la invaginación avance de manera ordenada hasta formar una estructura con forma de cuello estrecho que permanece conectada a la superficie celular.
La separación definitiva de la vesícula respecto de la membrana plasmática constituye una de las etapas más críticas del proceso. Actualmente se sabe que esta escisión depende principalmente de la acción de la dinamina, una proteína con actividad de hidrólisis de trifosfato de guanosina. La dinamina se ensambla alrededor del cuello de la vesícula en formación y, mediante cambios conformacionales impulsados por energía química, provoca el estrangulamiento progresivo de la membrana hasta producir su separación completa.
Tras la escisión, la vesícula recién formada pierde rápidamente su cubierta de clatrina mediante un proceso denominado desrevestimiento. Esta etapa resulta esencial porque permite que la vesícula se fusione posteriormente con compartimentos endosómicos intracelulares. El reciclaje de la clatrina y de las proteínas adaptadoras posibilita su reutilización en nuevos ciclos de endocitosis.
Las vesículas pinocíticas se fusionan posteriormente con endosomas tempranos, donde comienza la clasificación intracelular de los materiales internalizados. En estos compartimentos, los receptores pueden separarse de sus ligandos debido a cambios en el pH luminal. Los receptores suelen reciclarse hacia la membrana plasmática para ser reutilizados, mientras que muchas de las moléculas captadas son transportadas hacia lisosomas para su degradación o procesamiento.
La participación del ATP resulta indispensable durante múltiples etapas de la pinocitosis. La formación de vesículas, el ensamblaje y desensamblaje de complejos proteicos, la remodelación del citoesqueleto, el tráfico intracelular de vesículas y los mecanismos de reciclaje de membrana dependen de aportes continuos de energía metabólica. Por esta razón, la inhibición de la producción celular de ATP reduce significativamente la actividad endocítica.
El calcio extracelular también desempeña funciones reguladoras importantes. Los iones Ca²⁺ participan en numerosas vías de señalización intracelular que modulan la dinámica del citoesqueleto de actina, la actividad de proteínas motoras y la organización de complejos endocíticos. Aunque los primeros modelos fisiológicos proponían que el calcio actuaba directamente sobre filamentos contráctiles para facilitar la separación de las vesículas, investigaciones posteriores han demostrado que su función es considerablemente más compleja e involucra múltiples proteínas reguladoras que coordinan la formación, el movimiento y la maduración de las vesículas endocíticas.
La pinocitosis constituye un mecanismo esencial para la supervivencia celular. Participa en la adquisición de hierro mediante la internalización de transferrina, en la captación de colesterol transportado por lipoproteínas de baja densidad, en la regulación de receptores hormonales, en la presentación de antígenos por células inmunitarias y en la comunicación entre células mediante factores de crecimiento y citocinas. Alteraciones en cualquiera de estas etapas pueden contribuir al desarrollo de enfermedades metabólicas, neurodegenerativas, inmunológicas y neoplásicas.
Asimismo, numerosos virus, bacterias intracelulares y toxinas aprovechan las rutas pinocíticas para ingresar a las células huésped. Por esta razón, el estudio de la pinocitosis posee una enorme relevancia biomédica, ya que permite comprender tanto procesos fisiológicos normales como mecanismos patológicos asociados a infecciones y enfermedades humanas.


Fuente y lecturas recomendadas:
- Anderson, R. G. W., Brown, M. S., & Goldstein, J. L. (1977). Role of the coated endocytic vesicle in the uptake of receptor-bound low density lipoprotein in human fibroblasts. Cell, 10(3), 351–364. https://doi.org/10.1016/0092-8674(77)90022-8
- Conner, S. D., & Schmid, S. L. (2003). Regulated portals of entry into the cell. Nature, 422(6927), 37–44. https://doi.org/10.1038/nature01451
- Doherty, G. J., & McMahon, H. T. (2009). Mechanisms of endocytosis. Annual Review of Biochemistry, 78, 857–902. https://doi.org/10.1146/annurev.biochem.78.081307.110540
- Kaksonen, M., & Roux, A. (2018). Mechanisms of clathrin-mediated endocytosis. Nature Reviews Molecular Cell Biology, 19(5), 313–326. https://doi.org/10.1038/nrm.2017.132
- Mayor, S., & Pagano, R. E. (2007). Pathways of clathrin-independent endocytosis. Nature Reviews Molecular Cell Biology, 8(8), 603–612. https://doi.org/10.1038/nrm2216
- McMahon, H. T., & Boucrot, E. (2011). Molecular mechanism and physiological functions of clathrin-mediated endocytosis. Nature Reviews Molecular Cell Biology, 12(8), 517–533. https://doi.org/10.1038/nrm3151
- Mercer, J., Schelhaas, M., & Helenius, A. (2010). Virus entry by endocytosis. Annual Review of Biochemistry, 79, 803–833. https://doi.org/10.1146/annurev-biochem-060208-104626
- Mettlen, M., Chen, P. H., Srinivasan, S., Danuser, G., & Schmid, S. L. (2018). Regulation of clathrin-mediated endocytosis. Annual Review of Biochemistry, 87, 871–896. https://doi.org/10.1146/annurev-biochem-062917-012644
- Rizzoli, S. O. (2014). Synaptic vesicle recycling: Steps and principles. EMBO Journal, 33(8), 788–822. https://doi.org/10.1002/embj.201386357
- Takei, K., McPherson, P. S., Schmid, S. L., & De Camilli, P. (1995). Tubular membrane invaginations coated by dynamin rings are induced by GTP-γ-S in nerve terminals. Nature, 374(6518), 186–190. https://doi.org/10.1038/374186a0
Aprende administración paso a paso

ADMINISTRACION DESDE CERO