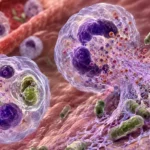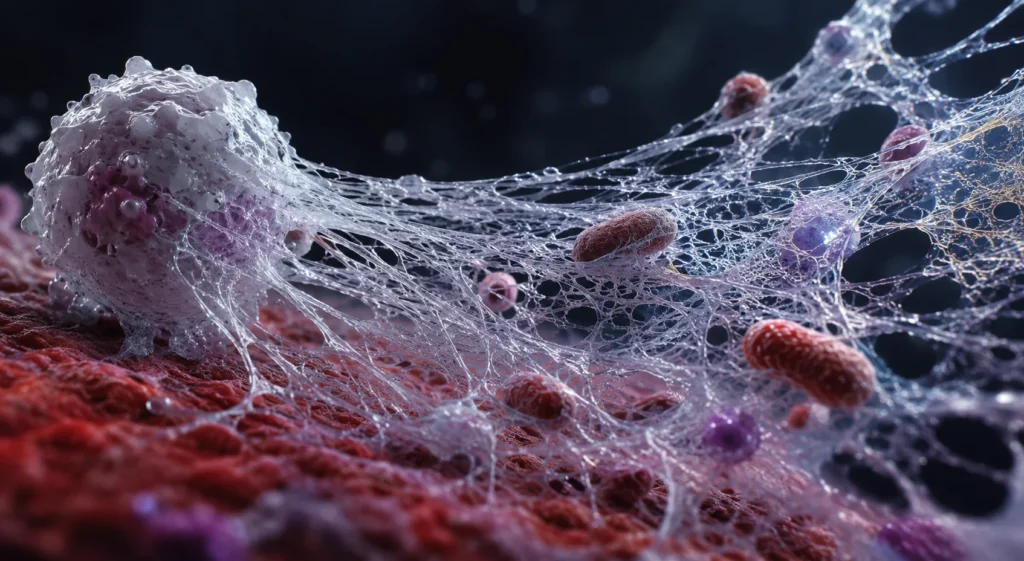
Las trampas extracelulares de neutrófilos, conocidas por las siglas NET (Neutrophil Extracellular Traps), constituyen un mecanismo altamente especializado de la inmunidad innata mediante el cual los neutrófilos inmovilizan y eliminan microorganismos extracelulares sin depender exclusivamente de la fagocitosis. Este proceso representa una estrategia complementaria a la degranulación y a la producción de especies reactivas del oxígeno, permitiendo contener la infección cuando la carga microbiana es elevada, cuando los microorganismos son demasiado grandes para ser fagocitados o cuando resulta esencial impedir su propagación hacia tejidos vecinos o al torrente sanguíneo. La formación de NET es una respuesta coordinada que integra señales provenientes del reconocimiento de patrones moleculares asociados con microorganismos, mediadores inflamatorios y moléculas derivadas del daño tisular, culminando con la expulsión al espacio extracelular de una red tridimensional de cromatina recubierta por proteínas antimicrobianas procedentes de los gránulos neutrofílicos. Esta organización convierte a la NET en una estructura con funciones tanto mecánicas como microbicidas.
La NET está constituida principalmente por ácido desoxirribonucleico descondensado proveniente del núcleo celular, aunque en determinadas circunstancias también puede originarse a partir del ácido desoxirribonucleico mitocondrial. Las largas fibras de cromatina actúan como un andamiaje sobre el cual permanecen firmemente adheridas numerosas proteínas con actividad antimicrobiana. Entre las más importantes destacan las histonas, la elastasa de neutrófilo, la mieloperoxidasa, la catepsina G, la proteinasa 3, la lactoferrina, las defensinas, la gelatinasa, la lisozima y diversas proteínas de los gránulos específicos y azurófilos. La combinación de estos componentes proporciona a la NET propiedades adhesivas, citotóxicas y antimicrobianas, permitiéndole capturar microorganismos y someterlos a una elevada concentración local de sustancias capaces de alterar su integridad estructural y funcional.
El proceso comienza cuando los neutrófilos reconocen microorganismos mediante receptores especializados de reconocimiento de patrones, como los receptores tipo Toll, receptores para el complemento, receptores Fc para inmunoglobulinas y otros sensores de moléculas derivadas de bacterias, hongos, virus y parásitos. Además de los productos microbianos, múltiples mediadores inflamatorios, entre ellos interleucina 8, factor activador de plaquetas, factor de necrosis tumoral alfa, interferones, productos del sistema del complemento y cristales endógenos, también pueden inducir la formación de NET. La activación simultánea de estos receptores desencadena complejas cascadas de señalización intracelular que modifican profundamente la organización del núcleo y de los gránulos citoplasmáticos.
Uno de los acontecimientos más importantes durante la formación de la NET es la pérdida de la compactación de la cromatina. En condiciones fisiológicas, el ácido desoxirribonucleico permanece estrechamente enrollado alrededor de las histonas, formando nucleosomas que mantienen el material genético altamente condensado. Durante la formación de la NET, esta organización cambia de manera radical gracias a la acción de diversas enzimas. La peptidil arginina deiminasa 4 cataliza la citrulinación de las histonas, convirtiendo residuos de arginina en citrulina. Esta modificación reduce la carga positiva de las histonas y disminuye su afinidad por el ácido desoxirribonucleico, favoreciendo la descondensación progresiva de la cromatina. Paralelamente, la elastasa de neutrófilo migra desde los gránulos hacia el núcleo, donde degrada parcialmente las histonas, potenciando aún más la relajación de la cromatina. La mieloperoxidasa también participa facilitando la reorganización nuclear mediante mecanismos que aún continúan investigándose.
En muchas formas de formación de NET, la producción de especies reactivas del oxígeno desempeña un papel esencial. La activación del complejo nicotinamida adenina dinucleótido fosfato oxidasa genera anión superóxido y otras especies reactivas que actúan como segundos mensajeros para activar las enzimas responsables de la descondensación cromatínica. Sin embargo, también se han descrito mecanismos independientes de este sistema oxidativo, lo que demuestra que existen diversas vías moleculares capaces de conducir a la liberación de NET dependiendo del estímulo y del contexto fisiopatológico.
Tradicionalmente se describió que la formación de NET culminaba con una modalidad específica de muerte celular denominada NETosis. Durante este proceso, desaparecen las membranas nucleares, la cromatina se mezcla con el contenido de los gránulos citoplasmáticos y finalmente la membrana plasmática se rompe, liberando al espacio extracelular la red de cromatina cubierta de proteínas antimicrobianas. Como consecuencia, el neutrófilo pierde su viabilidad y deja de participar en otras funciones inmunológicas.
Posteriormente se descubrió que no todas las NET requieren la muerte del neutrófilo. Existen mecanismos conocidos como NETosis vital, mediante los cuales determinadas porciones de cromatina son expulsadas en vesículas sin que ocurra destrucción inmediata de la membrana plasmática. En estos casos, el neutrófilo conserva su capacidad para desplazarse, fagocitar microorganismos y responder a nuevos estímulos inflamatorios. Incluso se ha demostrado que algunos neutrófilos pueden formar NET utilizando ácido desoxirribonucleico mitocondrial, preservando prácticamente intacto su núcleo.
La función más evidente de la NET consiste en inmovilizar microorganismos extracelulares. Las fibras de ácido desoxirribonucleico forman una compleja malla tridimensional cuya superficie posee abundantes cargas eléctricas y proteínas adhesivas que favorecen la unión física de bacterias, hongos, protozoarios y algunos virus. Esta captura mecánica impide que los microorganismos se dispersen por los tejidos o ingresen a la circulación sistémica, limitando así la extensión de la infección y concentrando el proceso inflamatorio en un sitio anatómico relativamente delimitado.
El atrapamiento físico tiene profundas implicaciones biológicas. Cuando los microorganismos permanecen confinados dentro de la red extracelular, aumenta considerablemente el tiempo de exposición a proteínas antimicrobianas presentes sobre la propia NET. En lugar de difundirse libremente por el tejido, las enzimas permanecen altamente concentradas alrededor del patógeno, creando un microambiente hostil que incrementa la eficacia de la respuesta inmunitaria.
Las histonas constituyen uno de los componentes antimicrobianos más importantes de la NET. Aunque normalmente participan en el empaquetamiento del ácido desoxirribonucleico nuclear, cuando son liberadas al espacio extracelular presentan una potente actividad citotóxica frente a numerosos microorganismos. Sus cargas positivas interactúan con las membranas celulares bacterianas, generalmente ricas en fosfolípidos con carga negativa, alterando su estabilidad, aumentando su permeabilidad y favoreciendo la muerte celular.
La elastasa de neutrófilo degrada proteínas estructurales y factores de virulencia producidos por numerosos microorganismos. Esta proteasa también destruye proteínas presentes en la superficie bacteriana necesarias para la adhesión, invasión tisular y evasión inmunológica. Al permanecer inmovilizada sobre la NET, la elastasa alcanza concentraciones locales muy superiores a las que existirían si simplemente fuera liberada al medio extracelular.
La mieloperoxidasa potencia el daño oxidativo mediante la generación de compuestos altamente reactivos derivados del peróxido de hidrógeno y de los haluros presentes en el medio extracelular. Estos productos oxidan proteínas, lípidos y ácidos nucleicos microbianos, comprometiendo múltiples funciones celulares indispensables para la supervivencia del microorganismo.
La lactoferrina contribuye al efecto antimicrobiano mediante el secuestro del hierro extracelular. Como numerosos microorganismos requieren hierro para sintetizar enzimas esenciales y multiplicarse, la disminución de su disponibilidad limita el crecimiento microbiano y potencia la acción de otros mecanismos de defensa presentes en la NET.
Las defensinas y otros péptidos antimicrobianos insertan poros en las membranas de bacterias y hongos, alterando el equilibrio osmótico y favoreciendo la lisis celular. La acción simultánea de proteasas, péptidos antimicrobianos, especies reactivas del oxígeno e histonas genera un efecto sinérgico considerablemente más eficaz que la acción aislada de cualquiera de estos componentes.
Las NET también favorecen indirectamente la fagocitosis. Al inmovilizar a los microorganismos, reducen su movilidad y los mantienen concentrados en un área limitada, facilitando que otros neutrófilos, macrófagos y células dendríticas entren en contacto con ellos. De esta manera, la red extracelular funciona como una plataforma de captura que incrementa la eficiencia de la eliminación posterior mediante fagocitosis. Además, muchos microorganismos atrapados experimentan daño parcial por las proteínas antimicrobianas, haciéndolos más susceptibles al reconocimiento y destrucción por las células fagocíticas.
Otra función relevante consiste en limitar la diseminación sistémica de los microorganismos. Al impedir que bacterias y hongos atraviesen el tejido conectivo o ingresen a vasos sanguíneos y linfáticos, las NET reducen la probabilidad de bacteriemia, fungemia y formación de focos infecciosos secundarios. Este efecto resulta especialmente importante durante las fases iniciales de una infección, cuando la respuesta inmunitaria adaptativa aún no se ha desarrollado plenamente.
Diversos microorganismos han evolucionado mecanismos específicos para escapar de las NET. Algunas bacterias producen desoxirribonucleasas extracelulares capaces de degradar la red de ácido desoxirribonucleico y recuperar su movilidad. Otras sintetizan proteínas que neutralizan histonas o inhiben la actividad de proteasas antimicrobianas. Ciertos hongos modifican la composición de su pared celular para disminuir la adhesión a la cromatina extracelular, mientras que algunos virus interfieren con las vías de señalización responsables de la formación de NET. Esta constante interacción representa un ejemplo de la coevolución entre los mecanismos defensivos del hospedador y las estrategias de evasión microbiana.
Aunque las NET desempeñan una función protectora fundamental, su producción excesiva o su eliminación ineficiente puede ocasionar daño tisular significativo. Las histonas extracelulares poseen marcada citotoxicidad sobre células endoteliales y epiteliales, favoreciendo lesión vascular y aumento de la permeabilidad capilar. Asimismo, la persistencia prolongada de redes de cromatina favorece la activación de la coagulación, la agregación plaquetaria y la formación de trombos, estableciendo una estrecha relación entre inmunidad e inmunotrombosis. También se ha demostrado que la exposición persistente de autoantígenos nucleares contenidos en las NET puede contribuir al desarrollo de enfermedades autoinmunes como el lupus eritematoso sistémico y algunas vasculitis asociadas con anticuerpos anticitoplasma de neutrófilos.
La eliminación fisiológica de las NET depende principalmente de desoxirribonucleasas plasmáticas y de la fagocitosis por macrófagos. Este proceso evita la acumulación excesiva de cromatina extracelular y limita la inflamación persistente. Cuando la degradación es insuficiente, las NET permanecen durante periodos prolongados en los tejidos, favoreciendo inflamación crónica, lesión endotelial y activación continua del sistema inmunitario.
Las trampas extracelulares de neutrófilos representan un sofisticado mecanismo efector de la inmunidad innata que combina propiedades mecánicas y bioquímicas. Su estructura de cromatina descondensada recubierta por histonas, enzimas proteolíticas y proteínas antimicrobianas permite inmovilizar microorganismos, impedir su diseminación, aumentar la eficacia de la fagocitosis y concentrar localmente múltiples mecanismos microbicidas. Sin embargo, el mismo potencial citotóxico que las convierte en una herramienta eficaz contra los patógenos puede transformarlas en un importante factor de daño tisular cuando su formación o eliminación pierde el equilibrio fisiológico.


Fuente y lecturas recomendadas:
- Brinkmann, V., Reichard, U., Goosmann, C., Fauler, B., Uhlemann, Y., Weiss, D. S., Weinrauch, Y., & Zychlinsky, A. (2004). Neutrophil extracellular traps kill bacteria. Science, 303(5663), 1532–1535. https://doi.org/10.1126/science.1092385
- Delgado-Rizo, V., Martínez-Guzmán, M. A., Iñiguez-Gutierrez, L., García-Orozco, A., Alvarado-Navarro, A., & Fafutis-Morris, M. (2017). Neutrophil extracellular traps and its implications in inflammation: An overview. Frontiers in Immunology, 8, 81. https://doi.org/10.3389/fimmu.2017.00081
- Jorch, S. K., & Kubes, P. (2017). An emerging role for neutrophil extracellular traps in noninfectious disease. Nature Medicine, 23(3), 279–287. https://doi.org/10.1038/nm.4294
- Papayannopoulos, V. (2018). Neutrophil extracellular traps in immunity and disease. Nature Reviews Immunology, 18(2), 134–147. https://doi.org/10.1038/nri.2017.105
- Pilsczek, F. H., Salina, D., Poon, K. K. H., Fahey, C., Yipp, B. G., Sibley, C. D., Robbins, S. M., Green, F. H. Y., Surette, M. G., Sugai, M., Bowden, M. G., Hussain, M., Zhang, K., & Kubes, P. (2010). A novel mechanism of rapid nuclear neutrophil extracellular trap formation in response to Staphylococcus aureus. Journal of Immunology, 185(12), 7413–7425. https://doi.org/10.4049/jimmunol.1000675
- Sorvillo, N., Cherpokova, D., Martinod, K., & Wagner, D. D. (2019). Extracellular DNA NET-works with dire consequences for health. Circulation Research, 125(4), 470–488. https://doi.org/10.1161/CIRCRESAHA.119.314581
- Thiam, H. R., Wong, S. L., Wagner, D. D., & Waterman, C. M. (2020). Cellular mechanisms of NETosis. Annual Review of Cell and Developmental Biology, 36, 191–218. https://doi.org/10.1146/annurev-cellbio-020520-111016
- Yipp, B. G., & Kubes, P. (2013). NETosis: How vital is it? Blood, 122(16), 2784–2794. https://doi.org/10.1182/blood-2013-04-457671
Aprende administración paso a paso

ADMINISTRACION DESDE CERO