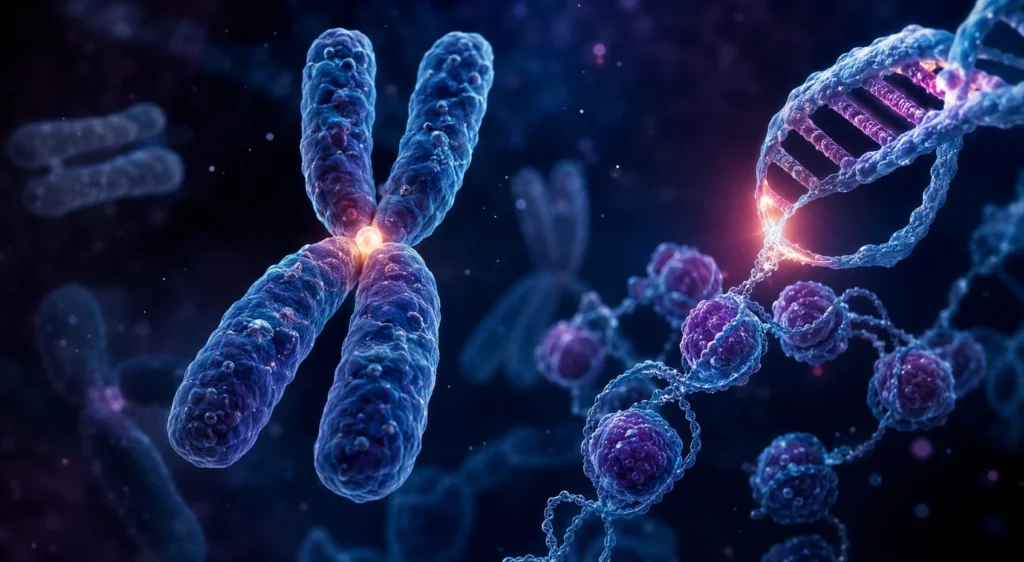
La replicación de los cromosomas constituye uno de los procesos más importantes para la continuidad de la vida, ya que permite que cada célula hija reciba una copia completa y funcional de la información genética contenida en el núcleo celular. Este proceso implica no solamente la duplicación de las moléculas de ácido desoxirribonucleico, sino también la reproducción de la compleja organización estructural que caracteriza a los cromosomas eucariotas. La replicación cromosómica es, por tanto, un fenómeno altamente coordinado que integra mecanismos de empaquetamiento, regulación genética, síntesis molecular, ensamblaje proteico y control del ciclo celular.
En las células humanas, el material genético se encuentra organizado en 46 cromosomas distribuidos en 23 pares. Veintidós de estos pares corresponden a autosomas y un par corresponde a los cromosomas sexuales. Cada par está constituido por cromosomas homólogos, uno heredado del progenitor masculino y otro heredado del progenitor femenino. Los cromosomas homólogos contienen genes localizados en posiciones equivalentes llamadas loci, aunque pueden presentar variantes diferentes de dichos genes. Esta organización permite mantener la estabilidad genética de la especie y al mismo tiempo favorecer la diversidad biológica.
Cada cromosoma está formado por una única molécula continua de ácido desoxirribonucleico asociada a numerosas proteínas estructurales y reguladoras. El ácido desoxirribonucleico humano contiene aproximadamente 3 200 millones de pares de bases distribuidos a lo largo de todo el genoma. Si esta molécula se encontrara extendida completamente alcanzaría una longitud cercana a los 2 metros, mientras que el diámetro del núcleo celular rara vez supera unas pocas micras. Esta enorme diferencia de escala hace indispensable la existencia de mecanismos extremadamente eficientes de compactación molecular.
La principal estrategia de empaquetamiento depende de las histonas. Estas proteínas poseen una elevada proporción de aminoácidos básicos, especialmente lisina y arginina, cuyas cargas positivas permiten una fuerte interacción electrostática con los grupos fosfato cargados negativamente del ácido desoxirribonucleico. Ocho moléculas de histonas forman complejos proteicos denominados octámeros de histonas. Alrededor de cada octámero se enrollan aproximadamente 147 pares de bases de ácido desoxirribonucleico formando una estructura denominada nucleosoma.
Los nucleosomas constituyen la unidad básica de organización de la cromatina. Miles de nucleosomas se distribuyen secuencialmente a lo largo de cada molécula de ácido desoxirribonucleico generando una fibra organizada que posteriormente experimenta niveles adicionales de plegamiento y condensación. Este empaquetamiento progresivo permite que enormes cantidades de información genética sean almacenadas dentro del núcleo sin comprometer la integridad estructural de la molécula.
Sin embargo, la función de las histonas no se limita al almacenamiento físico del material genético. El grado de compactación de la cromatina determina en gran medida la actividad funcional de los genes. Cuando el ácido desoxirribonucleico permanece fuertemente asociado a las histonas, las proteínas responsables de la transcripción encuentran dificultades para acceder a las secuencias reguladoras y codificantes. Como consecuencia, los genes contenidos en regiones altamente compactadas presentan una actividad transcripcional reducida o permanecen completamente silenciados.
Por el contrario, determinadas proteínas remodeladoras de cromatina pueden alterar la organización nucleosomal mediante procesos dependientes de energía química. Estas proteínas desplazan nucleosomas, modifican su posición o promueven cambios estructurales que exponen segmentos específicos del ácido desoxirribonucleico. De manera complementaria, diversas modificaciones químicas de las histonas, como acetilación, metilación, fosforilación y ubiquitinación, modifican la afinidad entre las histonas y el ácido desoxirribonucleico. Estas modificaciones generan regiones de cromatina más accesibles que permiten la unión de factores de transcripción y facilitan la síntesis de ácido ribonucleico.
Además de las histonas, los cromosomas contienen una gran variedad de proteínas no histonas. Algunas participan en la organización tridimensional del genoma formando dominios estructurales que mantienen la arquitectura nuclear. Otras actúan como factores de transcripción, represores, activadores, enzimas de reparación, proteínas de señalización o componentes de la maquinaria de replicación. Estas proteínas contribuyen a transformar al cromosoma en una estructura dinámica cuya actividad puede modificarse en respuesta a señales intracelulares y extracelulares.
La replicación cromosómica ocurre durante la fase de síntesis del ciclo celular. Antes de que una célula pueda dividirse, debe duplicar completamente su genoma. Este requisito garantiza que las dos células resultantes posean la misma información genética que la célula original. La duplicación comienza en regiones específicas denominadas orígenes de replicación. En estos sitios se ensamblan complejos proteicos especializados que reconocen secuencias particulares del ácido desoxirribonucleico y preparan el inicio del proceso replicativo.
La apertura inicial de la doble hélice es realizada por helicasas, enzimas capaces de romper los puentes de hidrógeno que mantienen unidas las bases complementarias. Como consecuencia se generan estructuras conocidas como horquillas de replicación. A medida que las helicasas avanzan, las dos cadenas parentales se separan progresivamente dejando expuestas sus secuencias nucleotídicas.
La separación de las cadenas genera tensiones mecánicas importantes en la molécula. Estas tensiones son aliviadas por enzimas denominadas topoisomerasas, capaces de cortar temporalmente una o ambas cadenas del ácido desoxirribonucleico para eliminar el superenrollamiento acumulado delante de las horquillas de replicación. Sin la actividad de estas enzimas, el avance de la maquinaria replicativa resultaría imposible.
Una vez expuestas las cadenas molde, proteínas estabilizadoras se unen al ácido desoxirribonucleico monocatenario para evitar su degradación o reasociación prematura. Posteriormente interviene un complejo enzimático denominado primasa, encargado de sintetizar pequeños fragmentos de ácido ribonucleico que sirven como puntos de inicio para la síntesis de nuevas cadenas.
Las polimerasas de ácido desoxirribonucleico incorporan nucleótidos complementarios siguiendo estrictamente las reglas de apareamiento de bases. La adenina se une con la timina y la citosina se une con la guanina. Gracias a esta complementariedad química, cada cadena parental funciona como molde para la generación de una nueva cadena complementaria.
La síntesis ocurre de manera diferente en cada una de las cadenas molde. Una de ellas permite una síntesis continua en dirección de avance de la horquilla de replicación. La otra requiere una síntesis discontinua mediante pequeños segmentos conocidos como fragmentos de Okazaki. Posteriormente estos fragmentos son unidos mediante la acción de ligasas, enzimas responsables de restaurar la continuidad química de la molécula.
La replicación del ácido desoxirribonucleico posee una extraordinaria precisión debido a la capacidad correctora de las polimerasas replicativas. Estas enzimas detectan errores de apareamiento y eliminan nucleótidos incorrectamente incorporados antes de continuar la síntesis. Adicionalmente, diversos sistemas de reparación posteriores a la replicación identifican y corrigen errores residuales. Gracias a estos mecanismos, la tasa de error final puede reducirse a aproximadamente una base incorrecta por cada miles de millones de nucleótidos incorporados.
El resultado de la replicación es la producción de dos moléculas hijas de ácido desoxirribonucleico genéticamente equivalentes. Cada una conserva una cadena parental y una cadena recién sintetizada. Este mecanismo recibe el nombre de replicación semiconservativa y constituye una de las propiedades fundamentales de la herencia genética.
La duplicación del ácido desoxirribonucleico es seguida casi inmediatamente por la reorganización de la cromatina. Las nuevas moléculas incorporan histonas recién sintetizadas y también reutilizan parte de las histonas preexistentes. Este proceso es esencial porque muchas modificaciones químicas presentes en las histonas participan en la transmisión de información epigenética. La distribución coordinada de histonas antiguas y nuevas contribuye a conservar patrones de expresión génica característicos de cada tipo celular.
Conforme avanza la replicación, proteínas ensambladoras especializadas depositan nucleosomas sobre el ácido desoxirribonucleico recién sintetizado. De esta manera se reconstruye gradualmente la arquitectura cromatínica. El proceso asegura que cada copia genética mantenga no solamente la secuencia de nucleótidos original, sino también una organización estructural compatible con la regulación adecuada de los genes.
Una vez completada la replicación, cada cromosoma ya no está formado por una sola molécula de ácido desoxirribonucleico, sino por dos moléculas idénticas estrechamente asociadas. Estas estructuras reciben el nombre de cromátidas hermanas. Cada cromátida contiene una copia completa de toda la información genética presente en el cromosoma original.
Las cromátidas hermanas permanecen unidas mediante complejos proteicos especializados localizados principalmente en la región denominada centrómero. Esta región cromosómica posee secuencias repetitivas de ácido desoxirribonucleico y proteínas específicas que permiten el ensamblaje del cinetocoro. El cinetocoro constituye una estructura multiproteica que servirá posteriormente como punto de unión para las fibras del huso mitótico.
La cohesión entre cromátidas hermanas resulta indispensable para garantizar una segregación cromosómica correcta. Mientras las cromátidas permanezcan unidas, el sistema celular puede verificar que todos los cromosomas estén adecuadamente conectados al huso mitótico. Cuando estas condiciones se cumplen, proteínas reguladoras activan mecanismos que rompen de forma controlada los complejos de cohesión.
Durante la mitosis, las fibras del huso ejercen fuerzas opuestas sobre cada cromátida hermana. Como resultado, ambas estructuras se separan y migran hacia polos opuestos de la célula. Una vez completada esta separación, cada cromátida pasa a considerarse un cromosoma independiente. Finalmente, la división celular distribuye los cromosomas entre las dos células hijas.
La correcta replicación cromosómica es indispensable para la estabilidad genética. Alteraciones en cualquiera de sus etapas pueden producir mutaciones, deleciones, duplicaciones, translocaciones o aneuploidías. Estos defectos pueden afectar la viabilidad celular, favorecer procesos de envejecimiento, originar enfermedades hereditarias o contribuir al desarrollo de diversos tipos de cáncer.
Por tanto, la replicación de los cromosomas representa un fenómeno biológico extraordinariamente complejo que integra la duplicación precisa del ácido desoxirribonucleico, la reorganización de la cromatina, la participación coordinada de histonas y proteínas no histonas, la formación de cromátidas hermanas y los mecanismos que aseguran una distribución equitativa del material genético. Gracias a esta coordinación molecular altamente regulada, cada generación celular conserva la información genética necesaria para mantener la estructura, función y continuidad de los organismos multicelulares.


Fuente y lecturas recomendadas:
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2022). Molecular Biology of the Cell (7th ed.). W. W. Norton & Company.
- Bell, S. P., & Labib, K. (2016). Chromosome duplication in Saccharomyces cerevisiae. Genetics, 203(3), 1027–1067.
- Burgers, P. M. J., & Kunkel, T. A. (2017). Eukaryotic DNA replication fork. Annual Review of Biochemistry, 86, 417–438.
- Clapier, C. R., Iwasa, J., Cairns, B. R., & Peterson, C. L. (2017). Mechanisms of action and regulation of ATP-dependent chromatin-remodelling complexes. Nature Reviews Molecular Cell Biology, 18(7), 407–422.
- Kornberg, R. D., & Lorch, Y. (2020). Primary role of the nucleosome. Molecular Cell, 79(3), 371–375.
- Luger, K., Mäder, A. W., Richmond, R. K., Sargent, D. F., & Richmond, T. J. (1997). Crystal structure of the nucleosome core particle at 2.8 Å resolution. Nature, 389(6648), 251–260.
- Meselson, M., & Stahl, F. W. (1958). The replication of DNA in Escherichia coli. Proceedings of the National Academy of Sciences, 44(7), 671–682.
- Peterson, C. L., & Almouzni, G. (2013). Nucleosome dynamics as modular systems that integrate DNA damage and repair. Cold Spring Harbor Perspectives in Biology, 5(9), a012658.
- Probst, A. V., Dunleavy, E., & Almouzni, G. (2009). Epigenetic inheritance during the cell cycle. Nature Reviews Molecular Cell Biology, 10(3), 192–206.
- Stillman, B. (2018). Histone modifications: Insights into their influence on gene expression. Cell, 175(1), 6–9.
- Yadav, T., Quivy, J. P., & Almouzni, G. (2018). Chromatin plasticity: A versatile landscape that underlies cell fate and identity. Science, 361(6409), 1332–1336.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO