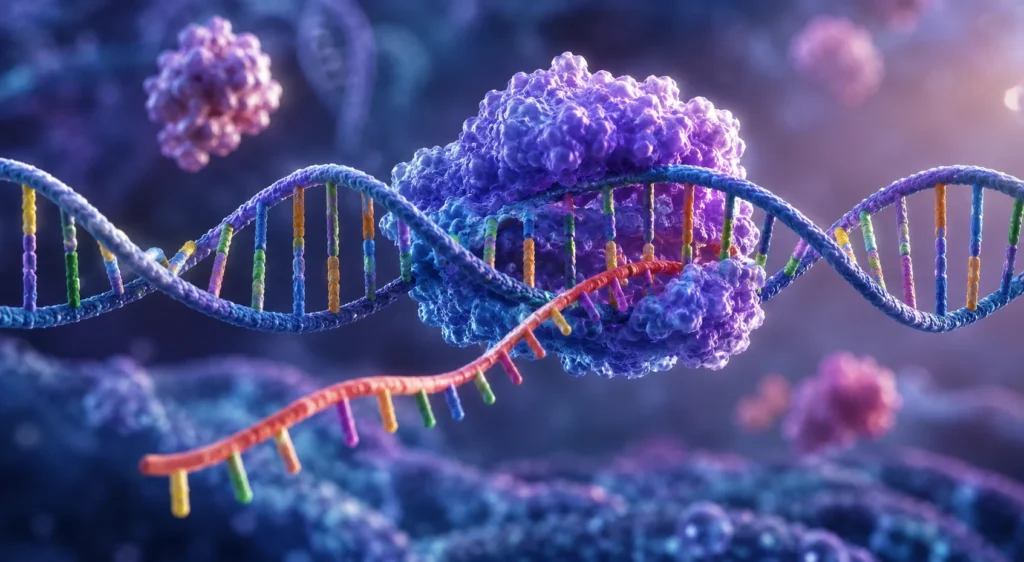
El montaje de la cadena de ARN constituye el proceso central de la transcripción genética, mediante el cual la información almacenada en la secuencia de nucleótidos del ADN es copiada de forma precisa hacia una molécula de ARN. Este mecanismo representa el primer paso en la expresión génica y es indispensable para la síntesis de proteínas, la regulación genética y el funcionamiento general de todas las células vivas. La transcripción se encuentra altamente conservada desde organismos procariotas hasta organismos eucariotas debido a que permite convertir la información genética estable del ADN en moléculas funcionales capaces de participar en numerosos procesos celulares. La síntesis de ARN es catalizada por una enzima especializada denominada ARN polimerasa, una proteína compleja capaz de reconocer secuencias específicas del ADN, separar temporalmente las hebras de la doble hélice, seleccionar nucleótidos complementarios y catalizar la formación de enlaces fosfodiéster que generan una nueva cadena de ARN.
La ARN polimerasa es una de las enzimas más sofisticadas presentes en la célula. Su estructura tridimensional contiene múltiples dominios funcionales responsables del reconocimiento del ADN, la apertura de la doble hélice, la selección de nucleótidos, la catálisis química de la polimerización y la corrección parcial de errores. En organismos eucariotas existen varias ARN polimerasas especializadas. La ARN polimerasa I sintetiza la mayor parte del ARN ribosómico, la ARN polimerasa II produce los ARN mensajeros y numerosos ARN reguladores, mientras que la ARN polimerasa III sintetiza ARN de transferencia y otros ARN pequeños. Cada una de estas enzimas presenta adaptaciones estructurales que les permiten reconocer conjuntos específicos de genes y participar en distintos procesos regulatorios.
El inicio de la transcripción depende del reconocimiento de regiones específicas del ADN denominadas promotores. Estas secuencias constituyen señales moleculares que indican dónde debe comenzar la síntesis del ARN. El promotor contiene motivos nucleotídicos característicos capaces de ser reconocidos por proteínas reguladoras y por la ARN polimerasa. En bacterias, la identificación del promotor es facilitada por factores sigma que dirigen la enzima hacia secuencias específicas. En células eucariotas intervienen numerosos factores generales de transcripción que ensamblan un complejo de iniciación sobre el promotor antes de permitir la unión estable de la ARN polimerasa.
La existencia de promotores es esencial porque garantiza que la transcripción se inicie en el lugar correcto del genoma. Sin este sistema de reconocimiento, la ARN polimerasa podría comenzar la síntesis en posiciones aleatorias, generando moléculas de ARN defectuosas o carentes de función biológica. La interacción entre la ARN polimerasa y el promotor depende de múltiples contactos moleculares entre aminoácidos específicos de la enzima y las bases nitrogenadas expuestas en el ADN, permitiendo una identificación altamente selectiva de las regiones que deben transcribirse.
Una vez que la ARN polimerasa se une al promotor, ocurre la formación del denominado complejo abierto de transcripción. Durante esta etapa la enzima induce el desenrollamiento local de la doble hélice del ADN. Generalmente se separan entre 12 y 17 pares de bases, generando una región denominada burbuja de transcripción. Esta apertura es indispensable porque las bases nitrogenadas que servirán como molde para la síntesis del ARN deben quedar expuestas y accesibles para el apareamiento complementario con los nucleótidos entrantes.
La separación temporal de las dos hebras del ADN implica la ruptura de numerosos enlaces de hidrógeno que normalmente mantienen unidas ambas cadenas. Aunque cada enlace de hidrógeno individual es relativamente débil, la suma de miles de estos enlaces proporciona una considerable estabilidad a la molécula de ADN. La ARN polimerasa utiliza la energía derivada de sus interacciones con el ADN y de los cambios conformacionales de su estructura para mantener abierta la burbuja de transcripción durante el proceso de síntesis.
Tras la formación del complejo abierto, la ARN polimerasa inicia la elongación de la cadena de ARN. La enzima avanza a lo largo de una sola hebra del ADN, denominada hebra molde o hebra plantilla, desplazándose en dirección 3’→5’ sobre dicha hebra. Simultáneamente sintetiza el ARN en dirección 5’→3’. Esta orientación es una consecuencia directa de la química de los ácidos nucleicos y de la forma en que ocurre la reacción catalizada por la ARN polimerasa.
Durante la elongación, cada nucleótido de ARN es incorporado siguiendo estrictamente las reglas de complementariedad de bases. Cuando la hebra molde contiene una guanina, se incorpora una citosina en el ARN. Cuando contiene una citosina, se incorpora una guanina. Cuando aparece una adenina en el ADN, se incorpora un uracilo en el ARN. Finalmente, cuando la plantilla contiene una timina, se incorpora una adenina.
Base en ADN | Base complementaria en ARN |
Guanina | Citosina |
Citosina | Guanina |
Adenina | Uracilo |
Timina | Adenina |
Este apareamiento complementario constituye el fundamento molecular de la transferencia de información genética. Gracias a esta correspondencia específica, la secuencia de nucleótidos presente en el ADN puede copiarse con una elevada fidelidad hacia el ARN. Como resultado, la información genética almacenada en el genoma se conserva durante el proceso de transcripción y puede ser utilizada posteriormente en la síntesis proteica.
La incorporación de cada nucleótido comienza cuando un ribonucleósido trifosfato libre difunde hasta el sitio activo de la ARN polimerasa. Los principales sustratos utilizados son ATP, GTP, CTP y UTP. Estos nucleótidos se consideran activados porque contienen tres grupos fosfato unidos mediante enlaces fosfoanhidro de alta energía. La presencia de estos enlaces convierte a los nucleótidos trifosfato en moléculas capaces de aportar la energía necesaria para la polimerización.
Inicialmente, el nucleótido entrante establece puentes de hidrógeno con la base complementaria presente en la hebra molde de ADN. Este reconocimiento molecular asegura que solamente los nucleótidos correctos permanezcan suficientemente estables dentro del sitio activo de la enzima para participar en la reacción química posterior. La ARN polimerasa posee mecanismos estructurales que favorecen la incorporación de nucleótidos correctamente apareados y reducen la frecuencia de errores.
Una vez seleccionado el nucleótido apropiado, ocurre la reacción química fundamental de la transcripción. El grupo hidroxilo situado en el carbono 3’ de la ribosa correspondiente al último nucleótido incorporado ataca al fosfato alfa del nucleótido trifosfato entrante. Como consecuencia, se forma un nuevo enlace fosfodiéster que une ambos nucleótidos dentro de la cadena de ARN en crecimiento. Simultáneamente, los fosfatos beta y gamma son liberados en forma de pirofosfato.
La posterior hidrólisis del pirofosfato libera una cantidad adicional de energía que vuelve termodinámicamente irreversible la reacción de polimerización. Este mecanismo explica por qué la ARN polimerasa puede sintetizar cadenas de ARN largas y estables sin requerir una fuente energética adicional independiente. La energía necesaria para la formación de los enlaces fosfodiéster ya se encuentra almacenada en los nucleótidos trifosfato utilizados como sustratos.
A medida que la ARN polimerasa avanza sobre el ADN, mantiene una burbuja de transcripción móvil. En la región frontal se desenrolla una nueva porción de ADN, mientras que detrás de la enzima las dos hebras de ADN vuelven a aparearse espontáneamente. Este reapareamiento es favorecido por la elevada complementariedad y afinidad existente entre ambas cadenas de ADN.
La molécula de ARN recién sintetizada permanece asociada transitoriamente con la hebra molde mediante un corto híbrido ADN-ARN compuesto normalmente por aproximadamente 8 a 10 pares de bases. Sin embargo, conforme la ARN polimerasa continúa avanzando, los enlaces de hidrógeno entre el ARN y la plantilla se rompen progresivamente. Debido a que la interacción entre las dos hebras originales de ADN es más estable y extensa, estas vuelven a asociarse entre sí, desplazando al ARN naciente hacia el nucleoplasma.

La separación del ARN respecto del ADN resulta esencial para permitir que la molécula recién sintetizada adopte estructuras secundarias propias y participe posteriormente en procesos biológicos específicos. En el caso de los ARN mensajeros, la liberación permite que estos sean procesados, transportados y finalmente utilizados como molde para la síntesis de proteínas.
La elongación continúa hasta que la ARN polimerasa encuentra una secuencia terminadora. Las secuencias de terminación contienen señales específicas que indican el final de la unidad de transcripción. Estas señales desencadenan cambios estructurales en la ARN polimerasa, en el ARN recién sintetizado o en proteínas accesorias que conducen a la finalización del proceso.
En organismos procariotas existen mecanismos de terminación dependientes e independientes de proteínas auxiliares. En células eucariotas, la terminación suele involucrar procesos más complejos asociados al procesamiento del ARN. En todos los casos, el resultado final es la liberación de la molécula de ARN completa, la separación de la ARN polimerasa del ADN y la restauración de la estructura bicatenaria original del ADN.
La reutilización de la ARN polimerasa después de la terminación constituye una ventaja biológica importante. Una misma molécula enzimática puede participar repetidamente en múltiples ciclos de transcripción, permitiendo la síntesis continua de numerosas copias de ARN a partir de distintos genes según las necesidades fisiológicas de la célula.
La extraordinaria precisión de la transcripción se debe a la combinación de varios mecanismos. El reconocimiento específico del promotor asegura el inicio correcto. La complementariedad de bases garantiza la transferencia fiel de la información genética. La selección estructural de nucleótidos por la ARN polimerasa reduce errores de incorporación. Finalmente, determinados mecanismos de corrección contribuyen a mantener una elevada fidelidad global. Gracias a estos procesos coordinados, la información genética almacenada en el ADN puede expresarse de manera eficiente y confiable, permitiendo el funcionamiento normal de todos los sistemas biológicos.

Fuente y lecturas recomendadas:
- Bai, L., Santangelo, T. J., & Wang, M. D. (2006). Single-molecule analysis of RNA polymerase transcription. Annual Review of Biophysics and Biomolecular Structure, 35, 343–360. https://doi.org/10.1146/annurev.biophys.35.040405.102024
- Cramer, P. (2019). Organization and regulation of gene transcription. Nature, 573(7772), 45–54. https://doi.org/10.1038/s41586-019-1517-4
- Feklistov, A., Sharon, B. D., Darst, S. A., & Gross, C. A. (2014). Bacterial sigma factors: A historical, structural, and genomic perspective. Annual Review of Microbiology, 68, 357–376. https://doi.org/10.1146/annurev-micro-092412-155737
- Jinek, M., Coyle, S. M., & Doudna, J. A. (2011). Coupled 5′ nucleotide recognition and processivity in Xrn1-mediated mRNA decay. Molecular Cell, 41(5), 600–608. https://doi.org/10.1016/j.molcel.2011.02.004
- Kornberg, R. D. (2007). The molecular basis of eukaryotic transcription. Proceedings of the National Academy of Sciences, 104(32), 12955–12961. https://doi.org/10.1073/pnas.0704138104
- Larson, M. H., Mooney, R. A., Peters, J. M., Windgassen, T., Nayak, D., Gross, C. A., Block, S. M., Greenleaf, W. J., Landick, R., & Weissman, J. S. (2014). A pause sequence enriched at translation start sites drives transcription dynamics in vivo. Science, 344(6187), 1042–1047. https://doi.org/10.1126/science.1251871
- Nudler, E. (2009). RNA polymerase active center: The molecular engine of transcription. Annual Review of Biochemistry, 78, 335–361. https://doi.org/10.1146/annurev.biochem.77.073107.184655
- Roeder, R. G. (2019). 50+ years of eukaryotic transcription: An expanding universe of factors and mechanisms. Nature Structural & Molecular Biology, 26(9), 783–791. https://doi.org/10.1038/s41594-019-0287-x
- Steitz, T. A. (2009). The structural changes of T7 RNA polymerase from transcription initiation to elongation. Current Opinion in Structural Biology, 19(6), 683–690. https://doi.org/10.1016/j.sbi.2009.09.010
- Vannini, A., & Cramer, P. (2012). Conservation between the RNA polymerase I, II, and III transcription initiation machineries. Molecular Cell, 45(4), 439–446. https://doi.org/10.1016/j.molcel.2012.01.023
- Watson, J. D., Baker, T. A., Bell, S. P., Gann, A., Levine, M., & Losick, R. (2014). Molecular Biology of the Gene (7th ed.). Pearson.
- Weixlbaumer, A., Leon, K., Landick, R., & Darst, S. A. (2013). Structural basis of transcriptional pausing in bacteria. Cell, 152(3), 431–441. https://doi.org/10.1016/j.cell.2012.12.020
Aprende administración paso a paso

ADMINISTRACION DESDE CERO