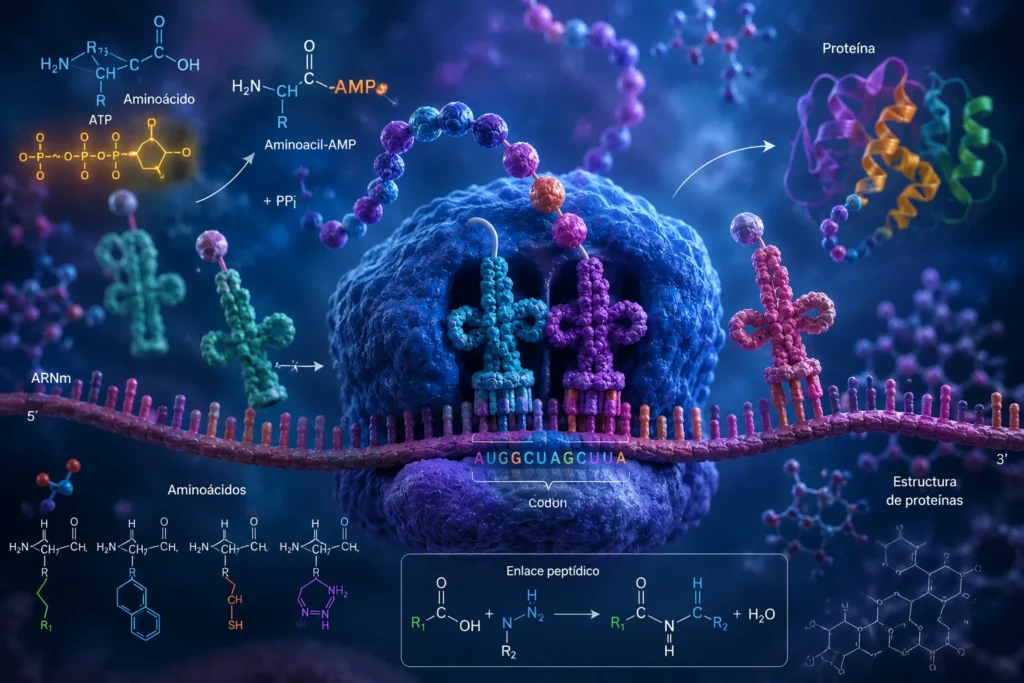
La síntesis proteica constituye uno de los procesos bioquímicos más complejos, regulados y energéticamente demandantes de la célula. Mediante este mecanismo, la información genética codificada en el ácido ribonucleico mensajero (ARNm) se traduce en una secuencia específica de aminoácidos que posteriormente adquirirá una estructura tridimensional funcional. La extraordinaria precisión de este proceso es indispensable para la supervivencia celular, ya que incluso errores mínimos en la incorporación de aminoácidos pueden alterar la estructura, estabilidad o actividad biológica de las proteínas. Por esta razón, la traducción proteica ha evolucionado incorporando múltiples mecanismos de reconocimiento molecular, corrección de errores y control energético que garantizan una fidelidad excepcional durante la síntesis de las cadenas polipeptídicas.
Activación del aminoácido
La primera etapa química de la síntesis proteica consiste en la activación de los aminoácidos. Antes de que un aminoácido pueda incorporarse a una proteína, debe ser energizado mediante una reacción catalizada por enzimas altamente específicas denominadas aminoacil-ARNt sintetasas.
Cada aminoácido posee una aminoacil-ARNt sintetasa específica o comparte una enzima con aminoácidos estructuralmente relacionados. Estas enzimas son responsables de reconocer simultáneamente un aminoácido determinado y el ARN de transferencia (ARNt) correspondiente. La precisión de este reconocimiento es fundamental para la fidelidad de la traducción, ya que el ribosoma no verifica directamente la identidad química del aminoácido transportado por el ARNt; depende por completo de la exactitud de la carga realizada por estas enzimas.
Durante la activación, el aminoácido reacciona con una molécula de ATP. Como resultado se forma un intermediario denominado aminoacil-adenilato o aminoácido-AMP. Simultáneamente, el ATP se hidroliza liberando pirofosfato (PPi). Posteriormente, el pirofosfato es degradado por una pirofosfatasa inorgánica en dos moléculas de fosfato inorgánico, lo que vuelve prácticamente irreversible la reacción.
Esta etapa consume el equivalente a dos enlaces fosfato de alta energía. La energía liberada durante la hidrólisis del ATP queda almacenada transitoriamente en el enlace formado entre el aminoácido y el AMP, constituyendo una reserva energética que será utilizada posteriormente durante la formación del enlace peptídico.
Las aminoacil-ARNt sintetasas poseen además mecanismos de corrección molecular denominados edición o proofreading. Estos sistemas permiten detectar y eliminar aminoácidos incorrectamente activados antes de que sean incorporados a la proteína. Gracias a estos mecanismos, la frecuencia de errores puede reducirse hasta aproximadamente un aminoácido incorrecto por cada 10.000 a 100.000 aminoácidos incorporados.
Formación del complejo aminoácido-ARNt
Una vez formado el aminoacil-adenilato, el aminoácido activado es transferido al extremo 3’ del ARNt correspondiente.
Todos los ARNt poseen una secuencia terminal conservada constituida por los nucleótidos citosina-citosina-adenina (CCA), localizada en el extremo aceptor. El grupo carboxilo del aminoácido se une covalentemente al grupo hidroxilo presente en la ribosa de la adenosina terminal.
Como resultado se forma un enlace éster de alta energía entre el aminoácido y el ARNt, generándose el complejo aminoacil-ARNt. Durante esta reacción se libera AMP.
El enlace éster formado representa un depósito de energía química. La energía originalmente obtenida de la hidrólisis del ATP permanece almacenada en dicho enlace y será utilizada posteriormente para impulsar la síntesis del enlace peptídico sin necesidad de un aporte energético directo adicional durante esa reacción específica.
Cada ARNt actúa como un adaptador molecular que conecta el lenguaje nucleotídico del ARNm con el lenguaje aminoacídico de las proteínas. De esta forma, los codones del ARNm pueden traducirse con precisión en una secuencia específica de aminoácidos.
La especificidad del sistema depende principalmente de las aminoacil-ARNt sintetasas, que reconocen características estructurales particulares tanto del aminoácido como del ARNt correspondiente. Este reconocimiento involucra secuencias específicas de nucleótidos, elementos tridimensionales del ARNt y determinantes estructurales distribuidos a lo largo de la molécula.
Reconocimiento del codón en el ribosoma
Una vez cargado, el aminoacil-ARNt es transportado hacia el ribosoma, la maquinaria molecular encargada de la síntesis proteica.
Los ribosomas están constituidos por una subunidad menor y una subunidad mayor formadas por ácido ribonucleico ribosómico (ARNr) y proteínas ribosómicas. La subunidad menor participa principalmente en la decodificación del mensaje genético, mientras que la subunidad mayor cataliza la formación de los enlaces peptídicos.
Durante la elongación, el aminoacil-ARNt ingresa inicialmente al sitio A del ribosoma. El anticodón presente en el ARNt establece apareamiento de bases con el codón complementario del ARNm mediante enlaces de hidrógeno.
Este reconocimiento constituye una etapa crítica para la fidelidad de la traducción. El ribosoma monitorea cuidadosamente la geometría del apareamiento codón-anticodón. Solamente cuando se establece una interacción correcta se desencadenan los cambios conformacionales necesarios para permitir la incorporación del aminoácido.
La precisión del reconocimiento depende tanto de la complementariedad de bases como de mecanismos cinéticos de selección molecular. Los complejos incorrectos presentan menor estabilidad y son rechazados antes de que ocurra la incorporación definitiva del aminoácido.
Factores de elongación dependientes de GTP participan activamente en esta etapa. Estos factores actúan como sistemas de verificación molecular que aumentan significativamente la exactitud del proceso traduccional.
Formación del enlace peptídico
Una vez que el aminoacil-ARNt ha sido correctamente posicionado en el sitio A, ocurre la reacción central de la síntesis proteica: la formación del enlace peptídico.
La catálisis de esta reacción es realizada por la actividad peptidil transferasa localizada en la subunidad mayor del ribosoma. De manera notable, esta actividad catalítica no depende principalmente de proteínas, sino del ARN ribosómico, lo que convierte al ribosoma en una ribozima.
La reacción implica el ataque nucleofílico del grupo amino libre del aminoácido recién incorporado sobre el grupo carbonilo del aminoácido terminal de la cadena polipeptídica unida al ARNt localizado en el sitio P.
Como consecuencia, la cadena polipeptídica completa es transferida desde el ARNt del sitio P hacia el aminoácido situado en el sitio A. El nuevo aminoácido pasa entonces a formar parte del extremo carboxilo terminal de la proteína en crecimiento.
La energía necesaria para esta reacción no procede directamente de ATP o GTP, sino de la energía almacenada previamente en el enlace éster de alta energía presente en el aminoacil-ARNt. Esto demuestra la importancia de la etapa inicial de activación de los aminoácidos.
Después de cada reacción de transferencia peptídica, la cadena polipeptídica aumenta su longitud en un residuo aminoacídico y queda preparada para recibir un nuevo aminoácido durante el siguiente ciclo de elongación.
Aporte energético para la elongación
La síntesis proteica requiere un consumo energético considerable debido a la necesidad de mantener simultáneamente velocidad, precisión y direccionalidad.
La carga inicial del aminoácido consume una molécula de ATP equivalente a dos enlaces fosfato de alta energía. Posteriormente, durante la elongación, la entrada del aminoacil-ARNt al sitio A requiere la hidrólisis de una molécula de GTP. Después de la formación del enlace peptídico, la translocación del ribosoma a lo largo del ARNm consume una segunda molécula de GTP.
Por consiguiente, para incorporar un único aminoácido a una proteína se requieren:
- Dos enlaces fosfato de alta energía derivados de la hidrólisis de ATP durante la activación del aminoácido.
- Un enlace fosfato de alta energía derivado de la hidrólisis de GTP durante la entrega del aminoacil-ARNt.
- Un enlace fosfato de alta energía derivado de la hidrólisis de GTP durante la translocación ribosómica.
El costo total corresponde a cuatro enlaces fosfato de alta energía por aminoácido incorporado.
Este elevado requerimiento energético refleja la enorme importancia biológica de la fidelidad traduccional. La célula invierte grandes cantidades de energía para minimizar errores que podrían generar proteínas defectuosas, agregados proteicos tóxicos o pérdida de funciones esenciales.
En organismos de crecimiento rápido, la síntesis proteica puede representar una de las mayores fracciones del gasto energético total celular, consumiendo una proporción significativa del ATP generado por el metabolismo.

Enlace peptídico
El enlace peptídico constituye la unión química fundamental que mantiene conectados los aminoácidos dentro de las proteínas.
Desde el punto de vista estructural, se trata de un enlace covalente formado entre el grupo carboxilo de un aminoácido y el grupo amino de otro aminoácido. Durante esta reacción se produce una condensación molecular en la que se elimina una molécula de agua.
Específicamente, el grupo hidroxilo procedente del grupo carboxilo de un aminoácido y un átomo de hidrógeno procedente del grupo amino del siguiente aminoácido se combinan para formar agua. Los átomos restantes establecen un nuevo enlace covalente carbono-nitrógeno denominado enlace peptídico.
El enlace peptídico presenta características electrónicas particulares debido a la resonancia entre el grupo carbonilo y el nitrógeno amídico. Como consecuencia, posee un carácter parcial de doble enlace que restringe la rotación libre alrededor del enlace carbono-nitrógeno.
Esta propiedad confiere rigidez estructural a las cadenas polipeptídicas y desempeña un papel esencial en el plegamiento tridimensional de las proteínas. La geometría relativamente plana de cada enlace peptídico contribuye a la formación de estructuras secundarias como las hélices α y las láminas β.
Durante la elongación, la formación repetitiva de enlaces peptídicos permite generar cadenas polipeptídicas progresivamente más largas. La secuencia exacta de aminoácidos queda determinada por la información codificada en el ARNm, mientras que las propiedades químicas de cada residuo aminoacídico determinarán posteriormente el plegamiento, estabilidad y función biológica de la proteína madura.
La síntesis proteica representa un extraordinario ejemplo de integración molecular en el que participan enzimas, ARN, ribosomas, factores proteicos y mecanismos energéticos altamente coordinados. La activación de aminoácidos, la carga de los ARNt, el reconocimiento codón-anticodón, la formación de enlaces peptídicos y la translocación ribosómica constituyen etapas sucesivas e interdependientes que permiten convertir la información genética en estructuras proteicas funcionales con una precisión cercana al límite físico de los sistemas biológicos.

Fuente y lecturas recomendadas:
- Ban, N., Beckmann, R., Cate, J. H. D., Dinman, J. D., Dragon, F., Ellis, S. R., Lafontaine, D. L. J., Lindahl, L., Liljas, A., Lipton, J. M., McAlear, M. A., Moore, P. B., Noller, H. F., Ortega, J., Panse, V. G., Ramakrishnan, V., Spahn, C. M. T., Steitz, T. A., Tchorzewski, M., … Warren, A. J. (2014). A new system for naming ribosomal proteins. Current Opinion in Structural Biology, 24, 165–169.
- Ibba, M., & Söll, D. (2000). Aminoacyl-tRNA synthesis. Annual Review of Biochemistry, 69, 617–650.
- Kapp, L. D., & Lorsch, J. R. (2004). The molecular mechanics of eukaryotic translation. Annual Review of Biochemistry, 73, 657–704.
- Ling, J., Reynolds, N., & Ibba, M. (2009). Aminoacyl-tRNA synthesis and translational quality control. Annual Review of Microbiology, 63, 61–78.
- Noller, H. F. (2005). RNA structure: Reading the ribosome. Science, 309(5740), 1508–1514.
- Rodnina, M. V. (2018). Translation in prokaryotes. Cold Spring Harbor Perspectives in Biology, 10(9), a032664.
- Schimmel, P. (2018). The emerging complexity of the tRNA world: Mammalian tRNAs beyond protein synthesis. Nature Reviews Molecular Cell Biology, 19(1), 45–58.
- Steitz, T. A. (2008). A structural understanding of the dynamic ribosome machine. Nature Reviews Molecular Cell Biology, 9(3), 242–253.
- Valle, M., Zavialov, A., Li, W., Stagg, S. M., Sengupta, J., Nielsen, R. C., Nissen, P., Harvey, S. C., Ehrenberg, M., & Frank, J. (2003). Incorporation of aminoacyl-tRNA into the ribosome as seen by cryo-electron microscopy. Nature Structural Biology, 10(11), 899–906.
- Voorhees, R. M., & Ramakrishnan, V. (2013). Structural basis of the translational elongation cycle. Annual Review of Biochemistry, 82, 203–236.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO