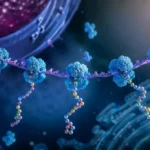
La síntesis de proteínas constituye uno de los procesos biológicos más importantes para la vida celular, ya que permite convertir la información genética almacenada en el ácido desoxirribonucleico en moléculas funcionales capaces de realizar prácticamente todas las actividades estructurales, metabólicas, reguladoras y defensivas de los organismos vivos. Este proceso ocurre mediante una secuencia ordenada de acontecimientos moleculares que culminan en la traducción de la información contenida en el ARN mensajero hacia una secuencia específica de aminoácidos. La maquinaria responsable de esta transformación es el ribosoma, un complejo ribonucleoproteico altamente especializado que actúa como centro catalítico de la síntesis proteica.
La traducción representa la etapa final de la expresión génica. Después de que la información genética ha sido copiada desde el ADN hacia una molécula de ARN mensajero mediante la transcripción, dicho ARN mensajero transporta la información codificada desde el núcleo hacia el citoplasma en las células eucariotas, o directamente hacia los ribosomas en las células procariotas. La secuencia de nucleótidos presente en el ARN mensajero determina de manera exacta el orden en que los aminoácidos serán incorporados a la proteína en formación, estableciendo así la estructura primaria de la molécula proteica y condicionando posteriormente su plegamiento tridimensional, estabilidad y función biológica.
El inicio de la traducción ocurre cuando una molécula de ARN mensajero entra en contacto con las subunidades ribosómicas. Este acontecimiento no sucede de forma aleatoria, sino que depende de señales específicas contenidas en la propia molécula de ARN mensajero. Los ribosomas deben identificar con precisión el punto exacto donde comenzará la síntesis proteica para garantizar que los aminoácidos sean incorporados en el orden correcto. Esta región corresponde al codón de inicio, que en la inmensa mayoría de los casos está constituido por la secuencia AUG. Dicho codón posee una importancia fundamental porque establece el marco de lectura que será utilizado durante toda la traducción. Una alteración en este punto provocaría que los tripletes posteriores fueran interpretados incorrectamente, originando proteínas defectuosas o completamente no funcionales.
La selección del codón de inicio es facilitada por numerosos factores proteicos especializados denominados factores de iniciación. Estas proteínas cooperan con las subunidades ribosómicas y con el ARN de transferencia iniciador para asegurar que la síntesis proteica comience exactamente en el sitio adecuado. En los organismos eucariotas, la subunidad pequeña del ribosoma reconoce inicialmente el extremo 5′ del ARN mensajero y posteriormente se desplaza a lo largo de la molécula hasta localizar el codón de inicio adecuado. Este mecanismo de exploración garantiza una elevada precisión en la identificación del punto de inicio de la traducción. La eficiencia de este reconocimiento está influenciada por secuencias nucleotídicas adyacentes que favorecen la correcta unión ribosómica y aumentan la fidelidad del proceso.
Una vez identificado el codón iniciador, se completa el ensamblaje del complejo de iniciación mediante la incorporación de la subunidad grande del ribosoma. En este momento queda constituido un complejo funcional capaz de comenzar la elongación de la cadena polipeptídica. El primer aminoácido incorporado suele ser la metionina en los eucariotas y la N-formilmetionina en los procariotas. Este aminoácido inicial marca el comienzo de la proteína naciente y desempeña un papel esencial en el establecimiento del proceso traduccional.
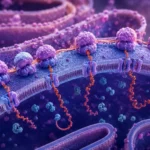
El ribosoma posee una arquitectura molecular extraordinariamente sofisticada. Está formado por ARN ribosómico y proteínas ribosómicas organizadas en dos subunidades funcionales. El ARN ribosómico no cumple únicamente funciones estructurales, sino que también constituye el principal componente catalítico responsable de la formación de los enlaces peptídicos. Este hallazgo transformó profundamente la comprensión de la biología molecular al demostrar que ciertas moléculas de ARN pueden actuar como enzimas, fenómeno conocido como actividad ribozímica.
Durante la elongación, el ribosoma se desplaza progresivamente a lo largo del ARN mensajero en dirección 5′→3′. Conforme avanza, va leyendo grupos sucesivos de tres nucleótidos denominados codones. Cada codón especifica un aminoácido concreto o una señal funcional dentro del proceso de traducción. El conjunto completo de correspondencias entre codones y aminoácidos constituye el código genético, uno de los sistemas de codificación biológica más conservados a lo largo de la evolución.
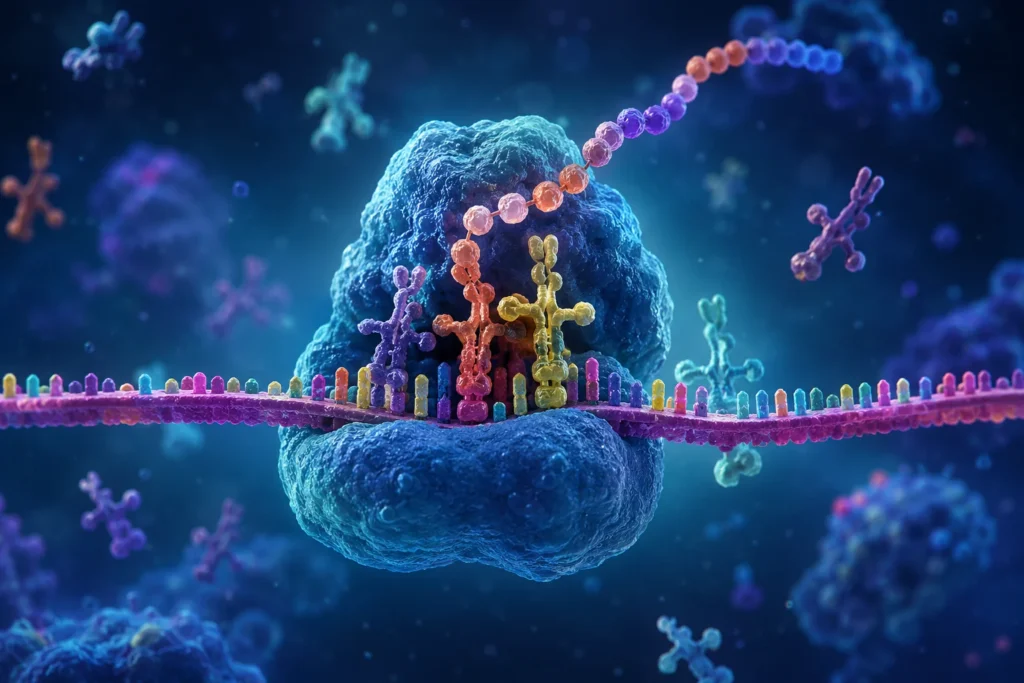
La lectura de los codones es posible gracias a la participación de los ARN de transferencia. Estas moléculas actúan como adaptadores moleculares que conectan el lenguaje de los ácidos nucleicos con el lenguaje de las proteínas. Cada ARN de transferencia posee un anticodón complementario a un codón específico del ARN mensajero y transporta simultáneamente el aminoácido correspondiente. Cuando el anticodón reconoce su codón complementario dentro del ribosoma, el aminoácido transportado queda posicionado adecuadamente para ser incorporado a la proteína en crecimiento.
La precisión de este reconocimiento es extraordinariamente elevada. Diversos mecanismos de control permiten detectar emparejamientos incorrectos entre codones y anticodones, minimizando la frecuencia de errores durante la síntesis proteica. Esta fidelidad resulta indispensable porque incluso pequeñas alteraciones en la secuencia aminoacídica pueden modificar significativamente la estructura y función de una proteína.
El ribosoma contiene tres sitios funcionales principales denominados sitio A, sitio P y sitio E. El sitio A recibe los ARN de transferencia cargados con aminoácidos. El sitio P mantiene el ARN de transferencia que sostiene la cadena polipeptídica en crecimiento. El sitio E constituye la vía de salida para los ARN de transferencia que ya han transferido su aminoácido. La coordinación entre estos tres sitios permite que la elongación ocurra de manera continua y altamente eficiente.
Cuando un nuevo ARN de transferencia ingresa al sitio A, el aminoácido que transporta se aproxima al extremo de la cadena polipeptídica ubicada en el sitio P. El centro peptidil transferasa del ribosoma cataliza entonces la formación de un enlace peptídico entre ambos aminoácidos. Como resultado, la cadena proteica se transfiere al ARN de transferencia situado en el sitio A. Posteriormente, el ribosoma experimenta un movimiento denominado translocación, mediante el cual avanza exactamente tres nucleótidos sobre el ARN mensajero. Este desplazamiento permite que el ARN de transferencia que contiene la cadena naciente pase al sitio P, mientras que el ARN de transferencia descargado se mueve al sitio E para abandonar el complejo ribosómico.
Este ciclo se repite una y otra vez durante toda la elongación. El ribosoma funciona de manera semejante a una máquina molecular programable que interpreta secuencialmente la información genética y la convierte en una cadena polipeptídica cuya composición está determinada exclusivamente por la secuencia de codones presente en el ARN mensajero. La velocidad de síntesis puede alcanzar varios aminoácidos por segundo en células eucariotas y decenas de aminoácidos por segundo en bacterias, permitiendo una producción proteica altamente eficiente.
Un aspecto notable de la traducción es la formación de polisomas o polirribosomas. Mientras un ribosoma avanza sobre una molécula de ARN mensajero, otros ribosomas pueden unirse al mismo transcrito y comenzar simultáneamente la síntesis de copias adicionales de la proteína. Este mecanismo incrementa enormemente la eficiencia de la expresión génica, permitiendo que una sola molécula de ARN mensajero produzca numerosas moléculas proteicas en un corto intervalo temporal.
La síntesis proteica continúa hasta que el ribosoma encuentra uno de los tres codones de terminación: UAA, UAG o UGA. Estos codones poseen una característica singular: no existen ARN de transferencia capaces de reconocerlos e incorporar aminoácidos. En consecuencia, cuando uno de estos tripletes alcanza el sitio A del ribosoma, la elongación se detiene.
La terminación requiere la acción de proteínas especializadas conocidas como factores de liberación. Estas moléculas reconocen específicamente los codones de terminación y desencadenan una serie de cambios estructurales dentro del ribosoma. Como consecuencia, se cataliza la hidrólisis del enlace que une la cadena polipeptídica al ARN de transferencia localizado en el sitio P. Esta reacción provoca la liberación de la proteína recién sintetizada hacia el citoplasma.
Una vez liberada, la proteína rara vez se encuentra inmediatamente lista para desempeñar su función biológica. La mayoría de las proteínas experimentan procesos posteriores de plegamiento, estabilización y modificación química. Chaperonas moleculares especializadas facilitan la adquisición de la conformación tridimensional correcta, mientras que diversas modificaciones postraduccionales pueden añadir grupos funcionales, fragmentar regiones específicas o dirigir la proteína hacia compartimentos celulares determinados. Estas etapas posteriores son esenciales para alcanzar la actividad biológica definitiva.
Tras la liberación de la proteína, las subunidades ribosómicas se disocian y quedan disponibles para participar en nuevos ciclos de traducción. Este reciclaje eficiente de los componentes celulares permite mantener una síntesis proteica continua y adaptada a las necesidades fisiológicas de la célula.
La extraordinaria precisión de la traducción es indispensable para la supervivencia celular. Cada proteína depende de una secuencia aminoacídica específica para adoptar la conformación tridimensional adecuada. Alteraciones en la fidelidad de la traducción pueden generar proteínas mal plegadas, agregados proteicos o pérdida de funciones celulares esenciales. Por esta razón, los sistemas traduccionales han evolucionado mecanismos de control de calidad altamente sofisticados que supervisan cada etapa del proceso, desde el reconocimiento del codón iniciador hasta la terminación de la síntesis.
La formación de proteínas en los ribosomas representa uno de los ejemplos más impresionantes de coordinación molecular en los sistemas biológicos. Mediante la interacción precisa entre ARN mensajero, ARN de transferencia, ARN ribosómico, proteínas ribosómicas y múltiples factores reguladores, la información genética puede transformarse en estructuras proteicas funcionales que sustentan todos los procesos fisiológicos de los organismos vivos.


Fuente y lecturas recomendadas:
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2022). Molecular biology of the cell (7th ed.). W. W. Norton & Company.
- Dever, T. E., Dinman, J. D., & Green, R. (2018). Translation elongation and recoding in eukaryotes. Cold Spring Harbor Perspectives in Biology, 10(8), a032649. https://doi.org/10.1101/cshperspect.a032649
- Green, R., & Noller, H. F. (1997). Ribosomes and translation. Annual Review of Biochemistry, 66, 679–716. https://doi.org/10.1146/annurev.biochem.66.1.679
- Hershey, J. W. B., Sonenberg, N., & Mathews, M. B. (2019). Principles of translational control. Cold Spring Harbor Perspectives in Biology, 11(9), a032607. https://doi.org/10.1101/cshperspect.a032607
- Lodish, H., Berk, A., Kaiser, C. A., Krieger, M., Bretscher, A., Ploegh, H., Amon, A., & Martin, K. C. (2021). Molecular cell biology (9th ed.). W. H. Freeman.
- Noller, H. F. (2005). RNA structure: Reading the ribosome. Science, 309(5740), 1508–1514. https://doi.org/10.1126/science.1112502
- Rodnina, M. V. (2018). Translation in prokaryotes. Cold Spring Harbor Perspectives in Biology, 10(9), a032664. https://doi.org/10.1101/cshperspect.a032664
- Steitz, T. A. (2008). A structural understanding of the dynamic ribosome machine. Nature Reviews Molecular Cell Biology, 9(3), 242–253. https://doi.org/10.1038/nrm2352
- Sonenberg, N., Hinnebusch, A. G., & Mathews, M. B. (Eds.). (2019). Translational control in biology and medicine (2nd ed.). Cold Spring Harbor Laboratory Press.
- Watson, J. D., Baker, T. A., Bell, S. P., Gann, A., Levine, M., & Losick, R. (2014). Molecular biology of the gene (7th ed.). Pearson.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO